La comunidad de Drosophila en las vegetaciones xerofíticas de la cuenca alta del río Paraná-Paraguay
Traducción automática
El artículo original está escrito en idioma EN (enlace para leerlo) .
Resumen
El género Drosophila es el más abundante en la familia Drosophilidae. Algunas especies son endémicas de ciertas regiones y otras son cosmopolitas. En Brasil, hay varios ecosistemas por explorar en cuanto a la composición y aspectos ecológicos de Drosophila. Las áreas xerofíticas son un ejemplo. Se pueden encontrar en el Sur y Sureste de Brasil como islas, resultado de cambios en los ciclos paleoclimáticos. El objetivo del presente trabajo es proporcionar información sobre la composición de la comunidad de Drosophila en ocho áreas xerofíticas (donde ocurre el cactus Cereus hildmaniannus) en el Sur y Sureste de Brasil. Este trabajo es un paso importante en el estudio de aspectos ecológicos cuantitativos de la comunidad de Drosophila en áreas xerofíticas que forman islas continentales en la región neotropical. La composición de la comunidad de Drosophila que se encontró es compatible con trabajos anteriores en varios aspectos. Los índices ecológicos mostraron una posible asociación positiva entre la diversidad y el grado de preservación de las áreas estudiadas. Las comunidades del estado de São Paulo presentaron la mayor similitud entre los sitios evaluados, aunque no se encontró una correlación estadísticamente significativa entre el índice de Jaccard y la distancia geográfica utilizando la prueba de Mantel.
Introducción
Aunque en el siglo XVIII Scopoli, Linné y Fabricius fueron los primeros en describir las especies que hoy se asignan a la familia Drosophilidae, Duda (1924a, 1924b, 1925) podría considerarse el pionero en la sistemática de este grupo. Además, Sturtevant, conocido por su trabajo clásico (1921, 1939, 1942), es uno de los investigadores más importantes dentro de esta familia. Los primeros datos sobre la comunidad de Drosophila brasileña fueron publicados por Duda (1925). En la región sureste de Brasil, Dobzhansky & Pavan (1943) fueron pioneros y describieron varias nuevas especies. Después de 1940, se publicó mucho trabajo sobre la fauna de Drosophila brasileña (Pavan & Cunha, 1947; Dobzhansky & Pavan, 1950; Pavan, 1950; 1959) incluyendo la primera lista de especies de Drosophila brasileñas (Mourão et al., 1965). Más tarde, otros autores analizaron la fauna de Drosophila en los dominios morfoclimáticos de Brasil en un esfuerzo por comprender mejor la distribución geográfica de las especies más comunes (Sene et al., 1980; Vilela et al., 1983; Val et al., 1981).
La familia Drosophilidae está compuesta por 65 géneros y más de 3,500 especies descritas que ocurren en una serie de ecosistemas en todo el mundo (Bächli, 1998). La mayoría de los géneros se encuentran en regiones tropicales. El Drosophila es el género más abundante y comprende alrededor del 53% del total de especies. Algunos de ellos son endémicos de ciertas regiones y otros son cosmopolitas, dispersos principalmente en asociación con la actividad humana. Grimaldi (1990) revisó la clasificación de la familia Drosophilidae basada en un análisis cladístico utilizando la máxima parsimonia. Este autor estudió 217 caracteres morfológicos de adultos y 120 especies representativas de la familia y concluyó que constituyen un grupo monofilético. El género Drosophila probablemente se originó en los trópicos (Stalker, 1976) y diferentes miembros de este expandieron su distribución a las zonas templadas, donde se seleccionó la capacidad de utilizar recursos en condiciones ambientales extremas. Este proceso podría haberse repetido varias veces durante la evolución del género.
Hoy en día, la mayoría de las investigaciones que utilizan especies de Drosophila se están llevando a cabo en los campos de la Biología Molecular y la Genética, pero fue solo a principios del siglo XX que Drosophila melanogaster fue reconocida como útil para análisis genéticos (Castle, 1906). A pesar del gran número de estudios que utilizan Drosophila como organismo modelo en estos campos, las especies brasileñas aún requieren más estudios sobre muchos aspectos de la ecología, sistemática, genética y evolución.
Se han realizado algunos estudios ecológicos con Drosophila, especialmente en lo que respecta a la actividad de las moscas durante el día (Klaczko et al., 1983; Medeiros, 2000), el período adecuado del día para el muestreo (Pavan et al., 1950; Belo & Oliveira-Filho, 1978), y la distribución espacial de las especies (horizontal: Dobzhanzky & Pavan, 1950; Burla et al., 1950; Pavan, 1959; Medeiros, 2000; y vertical: Sene et al., 1981; Kratz et al., 1982 e Tidon-Sklorz & Sene, 1992). Sin embargo, la comparación entre áreas ha recibido poca atención (Medeiros & Klaczko, 2004, por ejemplo) y sigue siendo un área de estudio abierta para las especies brasileñas de Drosophila.
Teniendo esto en cuenta, hay varios ecosistemas por explorar en Brasil en cuanto a la composición y aspectos ecológicos de Drosophila. Las áreas xerofíticas, determinadas por la presencia de vegetación de tipo seco con la ocurrencia de diferentes tipos de cactus, son un ejemplo de ello. Actualmente, así como en las áreas centrales de ocurrencia de cactus, como la Caatinga brasileña y el Chaco argentino, estas plantas se pueden encontrar en las regiones Sur, Sudeste y Central de Brasil en islas, resultado de los cambios paleoclimáticos cíclicos. Estos ciclos alternaron entre frío/seco y cálido/húmedo. En el primer caso, las áreas xerofíticas se expandieron desde las áreas centrales sobre el continente y en el segundo, se retrajeron formando refugios (islas) entre ellas (Bigarella et al., 1975; Ab’Saber, 1977; Vanzolini, 1981). Por lo tanto, los refugios son importantes para la biodiversidad, estudios ecológicos y evolutivos de varios grupos, incluyendo especies de Drosophila que se reproducen específicamente en tejido de cactus en descomposición, como lo hacen las moscas del grupo D. repleta.
El objetivo del presente trabajo es proporcionar información sobre la composición de la comunidad de Drosophila en ocho áreas xerofíticas (donde ocurre el cactus Cereus hildmaniannus) en las regiones sur y sureste de Brasil. Esto proporcionará información valiosa para futuros estudios sobre ecología, genética y la evolución de poblaciones de Drosophila geográficamente aisladas.
Material y métodos
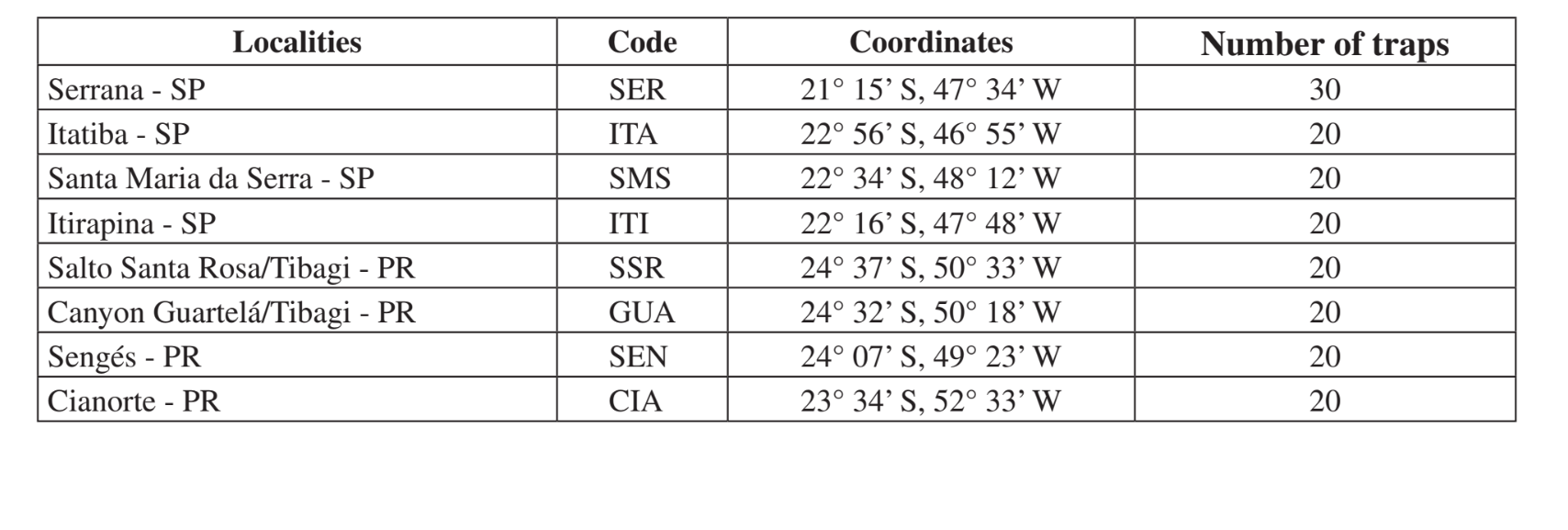
Los lugares y fechas de las colecciones se resumen en la Tabla 1. Las muestras fueron recolectadas en bosques compuestos de vegetación xerofítica en bosques mesofíticos en los valles de la porción superior de la cuenca del río Paraná-Paraguay. Estas áreas se encuentran en diferentes etapas de perturbación, que van desde el uso para la cría de ganado junto a una carretera (Itatiba - SP y Sengés - PR) hasta un área de preservación de Parque Estatal (Cañón Guartelá - PR). Todas las colecciones se realizaron al final del período húmedo, que corresponde a febrero a mayo en dos años diferentes (1999 y 2000).
Recolecciones
Durante tres días, las moscas adultas fueron atraídas por plátano y naranja fermentados en trampas abiertas, dispersas a no menos de 15 m entre sí, colgadas en ramas a ± 1.5 m de altura del suelo. Después de este período, las moscas fueron capturadas con redes entomológicas, puestas en frascos de vidrio con medio de cultivo y llevadas al laboratorio vivas, donde fueron identificadas.
Es difícil identificar a los miembros de los grupos Drosophila repleta y D. willistoni hasta cierto grado. Las hembras del primer grupo se muestran como no identificadas si la característica diagnóstica es el aedeago del macho. En términos del grupo Drosophila willistoni, la identificación utilizando rasgos genéticos es más confiable, pero no se realizó. Por lo tanto, las especies crípticas se agruparon como “grupo repleta” y “grupo D. willistoni”, respectivamente. Las especies no identificadas “tipo grupo repleta” no se consideraron en los análisis.
Análisis de datos
Para verificar la ocurrencia de especies cualitativamente, se utilizó el Método de Constancia de Ocurrencia (Dajoz, 1983). El valor de constancia (c) se obtuvo dividiendo el número de colecciones en las que ocurrió una especie por el número total de colecciones, y luego multiplicando ese resultado por 100. Las especies con índice c ≥ 50 se consideraron constantes. Las especies accesorias fueron aquellas con 25 ≤ c < 50. Las especies accidentales tenían c < 25. Las especies que ocurrieron en solo un área se consideraron exclusivas.
Las comunidades de Drosophila se analizaron utilizando varios índices ecológicos: Berger-Parker (1/d), Shannon-Wienner (H’), Simpson (D), Margalef (DMg), Menhinick (DMn) y Jaccard (J). Se utilizó un ANOVA no paramétrico, Kruskal-Wallis, para determinar si había diferencias en la abundancia mediana entre comunidades. Posteriormente, se realizó una comparación múltiple entre muestras (similar a la prueba de Tukey) para determinar si ocurrieron diferencias significativas entre las muestras (Zar, 1999).
Para verificar las relaciones entre comunidades, se realizaron dos análisis de clúster:
1) utilizando la presencia y ausencia de especies; y 2) utilizando el número de especímenes recolectados para cada especie en cada área. Los fenogramas se construyeron en el software Minitab® para Windows Release 10.1 seleccionando la distancia euclidiana y los métodos de enlace simple en ambos enfoques.
Para analizar si hay una asociación entre las similitudes de las comunidades y las distancias geográficas, se realizó una prueba de Mantel utilizando el software TFPGA (Miller, 1997) comparando dos matrices: índices de Jaccard y distancias geográficas entre un par de comunidades.
Resultados y discusión
En este estudio pudimos identificar un total de 10,464 moscas pertenecientes a 4 subgéneros, 10 grupos y al menos 30 especies diferentes de Drosophila (Tabla 2). Las especies constantes (c ≥ 50) representaron aproximadamente el 63% del total de especies recolectadas (19 de 30). Siete especies fueron consideradas accesorias (~ 23%) y se encontraron 4 especies accidentales (~ 13%). Todos los subgéneros mostraron especies constantes, excepto por el subgénero Dorsilopha que mostró solo una especie accidental (Drosophila busckii).
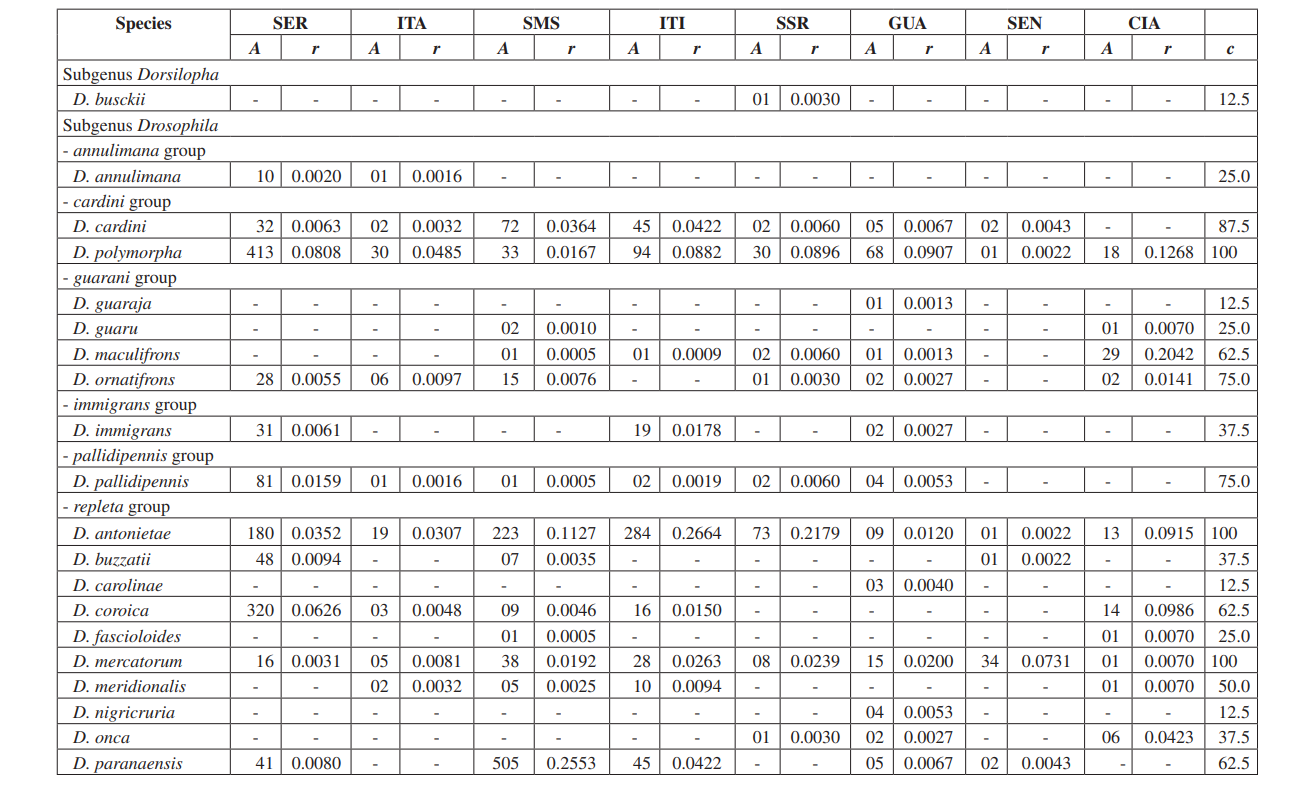
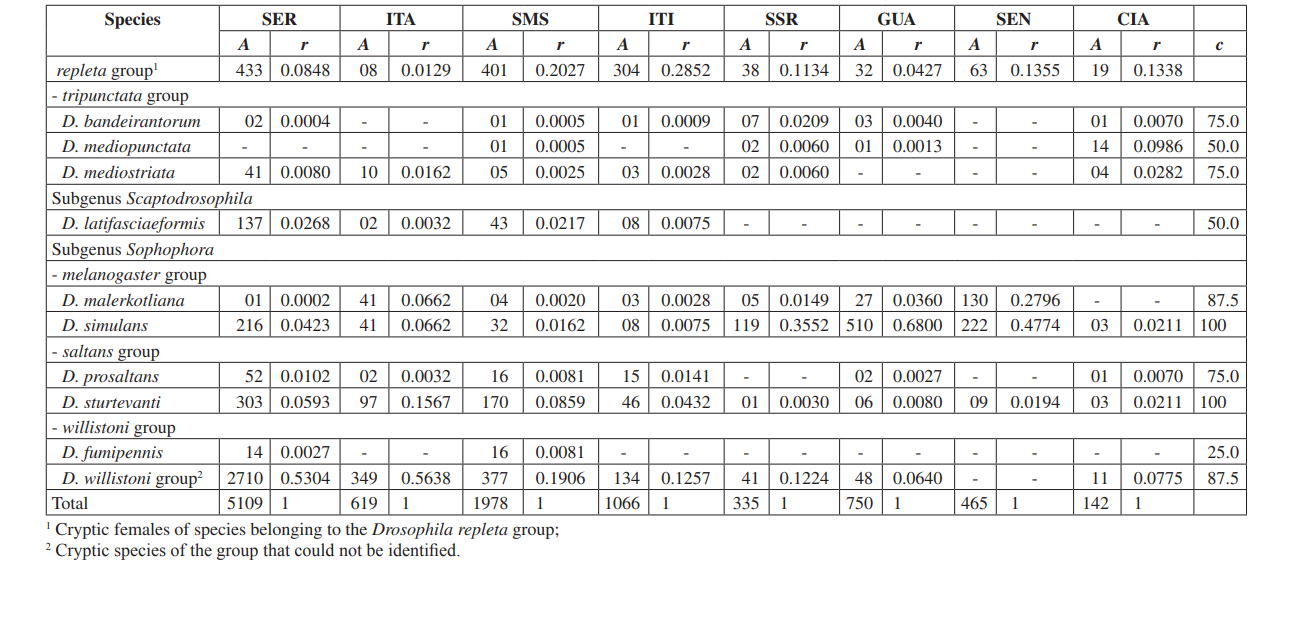
Se consideraron abundantes cinco especies (Drosophila polymorpha, D. antonietae, D. mercatorum, D. simulans y D. sturtevanti), y se recolectaron en las 8 áreas (Serrana - SP, Itatiba - SP, Santa Maria da Serra - SP, Itirapina - SP, Salto Santa Rosa - PR, Canyon Guartelá - PR, Sengés - PR y Cianorte - PR). Este resultado es compatible con encuestas previas de Drosophila.
Drosophila polymorpha se recolectó desde Guatemala hasta Brasil y en diferentes tipos de ambientes: bosques, cerrado, restinga, y también asociada con la presencia humana. Es una especie relativamente abundante en diferentes dominios morfoclimáticos, excepto en la caatinga (Sene et al., 1980). Se esperaba que Drosophila antonietae se recolectara en todas las áreas, ya que es una especie cactofílica que ocurre en las regiones sur y sureste de Brasil, y al norte del límite oriental del Chaco argentino. Esta especie siempre se encuentra asociada con el cactus columnar Cereus hildmaniannus en la vegetación xerofítica de galerías y bosques mesofíticos en los valles de la cuenca del río Paraná-Uruguai (Tidon-Sklorz & Sene, 2001). Este patrón de distribución que aísla a Drosophila antonietae de otras especies del grupo D. buzzatii coincide con patrones definidos de otros dípteros y también con regiones de endemismo, sugiriendo eventos de vicariancia como barreras zoogeográficas establecidas en eras geológicas anteriores o eventos climáticos que impidieron el flujo genético, permitiendo la acumulación de diferencias entre poblaciones (Amorin & Pires, 1996). Este patrón de distribución también se observa en relación con la distribución de plantas restringidas a las regiones xerofíticas de caatinga y chaco (Duvernell & Eanes, 2000).
Drosophila mercatorum es bastante común en ambientes naturales en América del Sur, especialmente en áreas abiertas (Sene et al., 1981; Vilela et al., 1983).
Drosophila simulans es una especie introducida que ha sido recolectada con alta frecuencia en varios lugares (Dobzhansky & Pavan, 1950; Pavan, 1959; Sene et al., 1980; Tidon-Sklorz & Sene, 1992), también en áreas abiertas (Perondini et al., 1979).
Drosophila sturtevanti es una especie ampliamente distribuida en América Central y del Sur que ocurre en diferentes dominios morfoclimáticos (Tidon-Sklorz & Sene, 1999).
Las especies exclusivas aparecieron en Salto Santa Rosa - PR (Drosophila busckii) y Canyon Guartelá - PR (Drosophila guaraja, D. carolinae y D. nigricruria). Drosophila busckii no se esperaba que fuera recolectada en Salto Santa Rosa - PR ya que es una especie introducida en Brasil y se asocia frecuentemente con ambientes modificados por el hombre (Tidon-Sklorz & Sene, 1999). Sin embargo, ha sido recolectada en ambientes naturales principalmente en vegetaciones secas abiertas (Bizzo & Sene, 1982; Tidon-Sklorz & Sene, 1992). Entre las especies exclusivas recolectadas en Canyon Guartelá - PR, Drosophila guaraja fue registrada en Brasil solo en Boracéia - SP (Val & Kaneshiro, 1988). Este es, por lo tanto, el primer registro de ella fuera de esta localidad en Brasil. Drosophila carolinae ha sido recolectada solo en selvas tropicales en Brasil (Vilela, 1983) y por esta razón no se esperaba encontrarla en áreas de vegetación xerofítica. Sin embargo, Canyon Guartelá - PR es una de las áreas más preservadas encuestadas y los cactus están distribuidos bajo una región boscosa dentro del Parque Estatal, un hecho que podría explicar su presencia. Drosophila nigricruria es la única especie exclusiva que tiene una amplia distribución geográfica. Sin embargo, fue recolectada solo en esta área en este estudio. Medeiros & Klaczko (2004) también recolectaron esta especie en la granja Barreiro Rico (entre 48° 05’ 11’’ O – 22° 41’ 15’’ S y 48° 04’ 52’’ O – 22° 41’ 26’’ S), un plateau interior que tiene un bosque semi caducifolio y uno de los tres bosques restantes en el Estado de São Paulo donde se realizaron colecciones por estos autores.
El subgénero Drosophila mostró la mayor riqueza (7 grupos y 22 especies), con 7 especies pertenecientes al grupo Drosophila repleta . La segunda mayor riqueza se encontró en el subgénero Sophophora (3 grupos y 6 especies). Esta distribución de riqueza entre subgéneros ha sido observada en otros estudios (Sene et al., 1981; Tidon-Sklorz & Sene, 1995; Tidon-Sklorz & Sene, 1999; Medeiros & Klaczko, 2004).
La especie de drosófilo introducida, Zaprionus indianus (Vilela, 1999), se encontró en todas las colecciones (1,459 especímenes en Serrana; 317 en Itatiba; 409 en Santa Maria da Serra; 302 en Itirapina; 42 en Salto Santa Rosa; 117 en Guartelá; 598 en Sengés; y 8 en Cianorte). Los eventos asociados a invasiones biológicas se dividen en tres categorías: llegada, establecimiento e integración (Vermeij, 1996). Según Moraes et al. (2000), Zaprionus indianus ha pasado por las tres y probablemente está compitiendo desfavorablemente con otras especies similares en el sureste de Brasil, ya que se ha observado una caída en la frecuencia relativa. La introducción de una especie exótica de Drosophila en Brasil no es un evento poco común. En 1976, Val & Sene (1980) recolectaron un gran número de Drosophila malerkotliana en el noreste de Brasil, una especie que no fue registrada en colecciones anteriores extensas. Actualmente, en nuestras encuestas, Drosophila malerkotliana se recolecta casi en todo Brasil, aunque siempre a una baja frecuencia relativa.
Veintitrés especies diferentes fueron recolectadas en Santa Maria da Serra - SP, 20 especies en Serrana - SP y en Canyon Guartelá - PR, 18 en Itirapina - SP, 17 en Cianorte - PR, 16 en Itatiba - SP y Salto Santa Rosa - PR y 9 en Sengés - PR (Tabla 3). Medeiros & Klaczko (2004) describieron el último inventario de la fauna de Drosophila en Brasil, encuestado en tres bosques remanentes en el Estado de São Paulo. Estos autores obtuvieron 57, 76 y 90 especies diferentes en las tres áreas estudiadas. Estos números de especies son mayores que los observados aquí, quizás como resultado de diferencias en el método de recolección o simplemente porque las áreas encuestadas son distintas en su constitución vegetal (bosque y vegetación xerofítica). La mayor abundancia total se detectó en Serrana - SP (4676 especímenes) y la más baja en Cianorte - PR (123 especímenes) (Tabla 3).
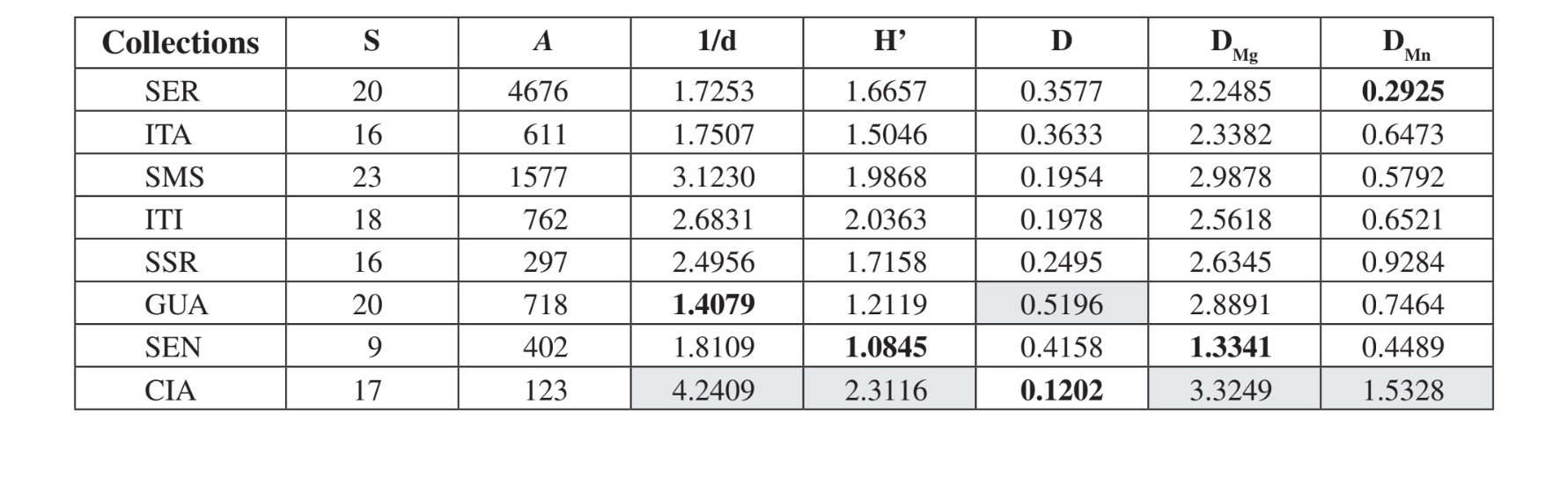
Según los índices ecológicos (Tabla 3), Cianorte - PR presentó valores más altos en casi todos los índices (1/d = 4.2409; H’ = 2.3116; DMg = 3.3249; DMn = 1.5328), excepto por Simpson (D) que mostró que Canyon Guartelá - PR es el área más
diversa (D = 0.5196). Canyon Guartelá - PR también mostró uno de los índices más altos en dos casos más (DMg = 2.8891; DMn = 0.7464). Sengés - PR, Itatiba - SP y Serrana - SP mostraron los valores más bajos para la mayoría de los índices. Santa Maria da Serra - SP, Itirapina - SP y Salto Santa Rosa - PR mostraron valores intermedios para la mayoría de los índices. Estas observaciones son compatibles con la evaluación superficial de la degradación en estas áreas. Cianorte - PR está ubicado junto a una carretera, pero en una ladera alta del río Ligeiro, con difícil acceso para la degradación humana. Canyon Guartelá - PR es un área boscosa dentro del Parque Estatal Canyon Guartelá del estado de Paraná, como se mencionó anteriormente. Por lo tanto, es un área altamente preservada. Sengés - PR es el área más degradada, junto a una carretera, hogar de la cría de ganado y bajo una alta influencia de la actividad humana. Serrana - SP es un área preservada en la cima de una colina que es difícil de acceder. Los índices de diversidad más bajos encontrados para esta área deben deberse al mayor número de trampas utilizadas en esta recolección (Tabla 1). Se esperaba que las otras áreas tuvieran valores intermedios de diversidad, ya que son pequeñas áreas de conservación dentro de granjas.
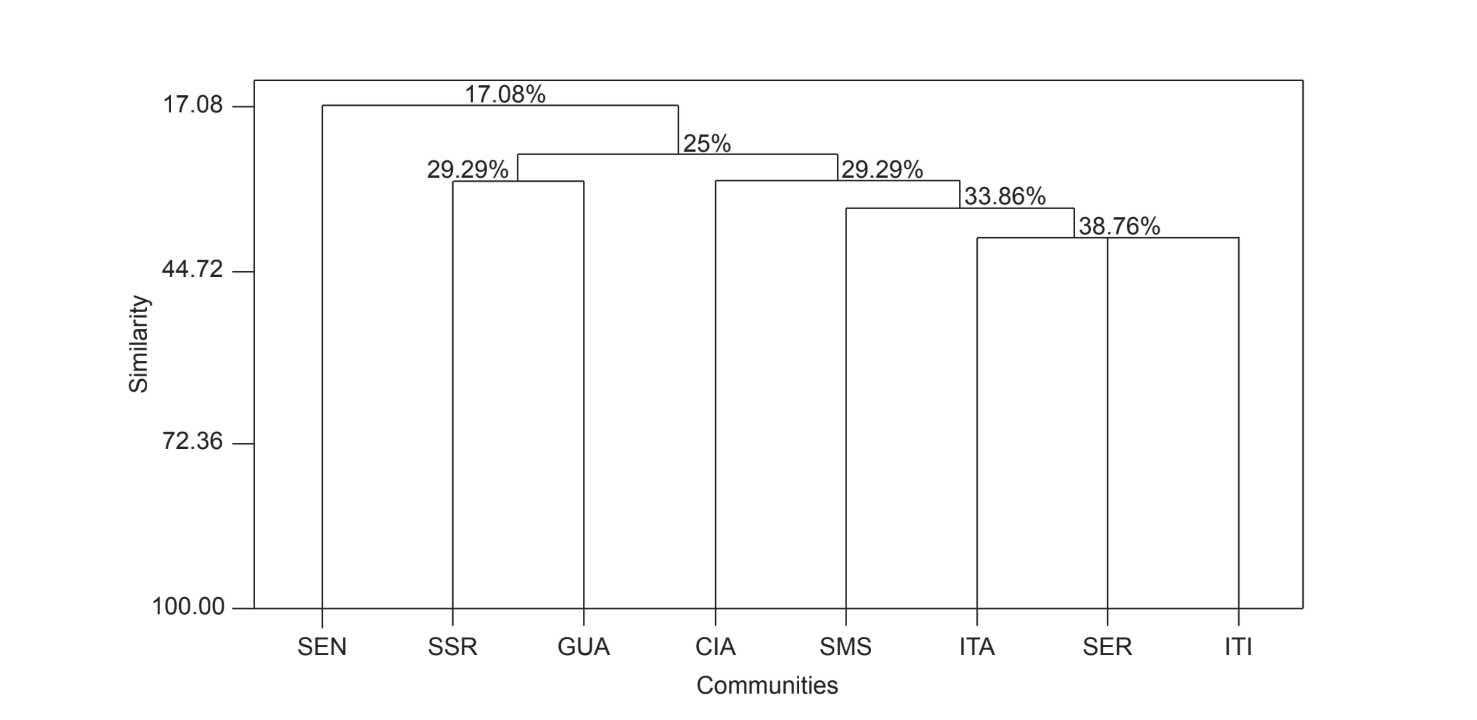
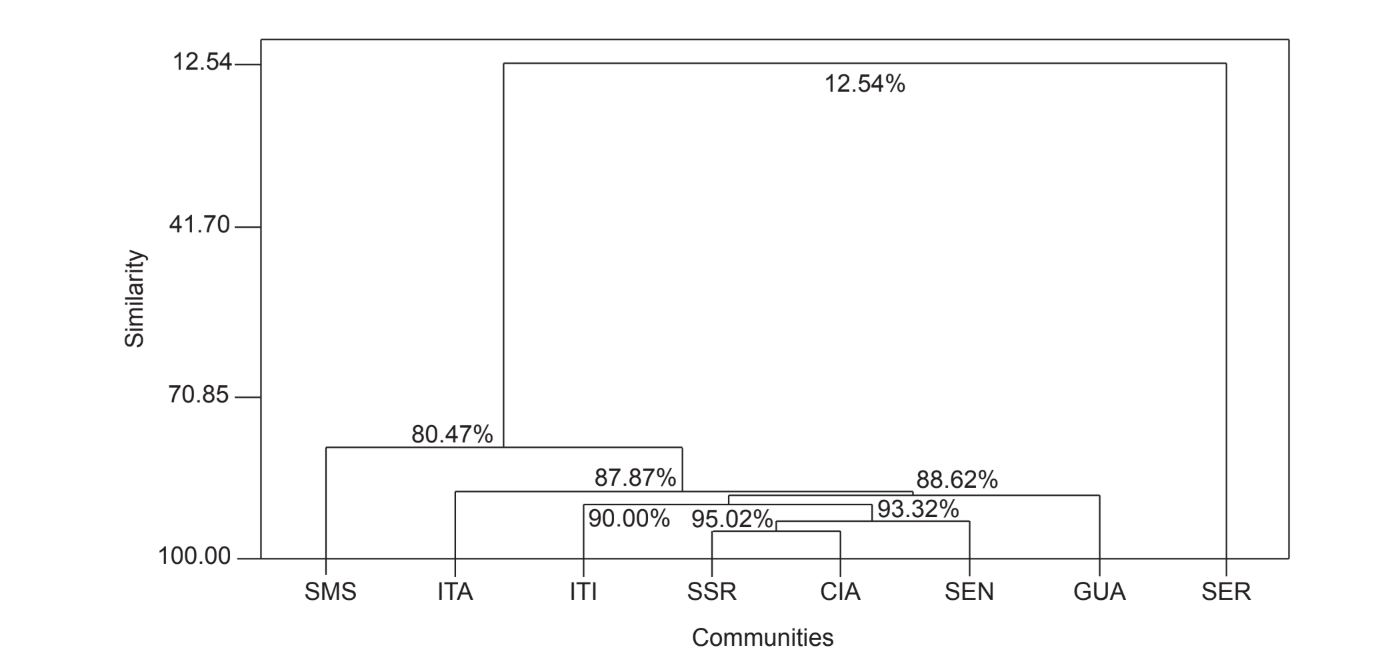
La comparación por pares de comunidades utilizando el índice de Jaccard (Tabla 4) mostró que las comunidades del estado de São Paulo (Serrana - SP, Itatiba - SP, Santa Maria da Serra - SP e Itirapina - SP) tenían la mayor similitud entre todas las comparaciones (J > 0.62). No se observó el mismo patrón entre las comunidades del estado de Paraná y también entre ambos estados, con la excepción del índice obtenido entre Santa Maria da Serra - SP y Cianorte - PR (J = 0.667), y entre Salto Santa Rosa - PR y Canyon Guartelá - PR (J = 0.636). Los análisis de agrupamiento utilizando datos de presencia y ausencia de especies (Fig. 1), a pesar de un bajo nivel de similitud (33.86%), también agruparon todas las comunidades del estado de São Paulo juntas. La comunidad más divergente fue Sengés - PR. La Fig. 2 representa el agrupamiento de todas las comunidades utilizando datos de abundancia absoluta (A). Se encontraron niveles más altos de similitud, pero no se encontró una asociación clara de comunidades. En esta figura, Serrana - SP fue la comunidad más diferente.
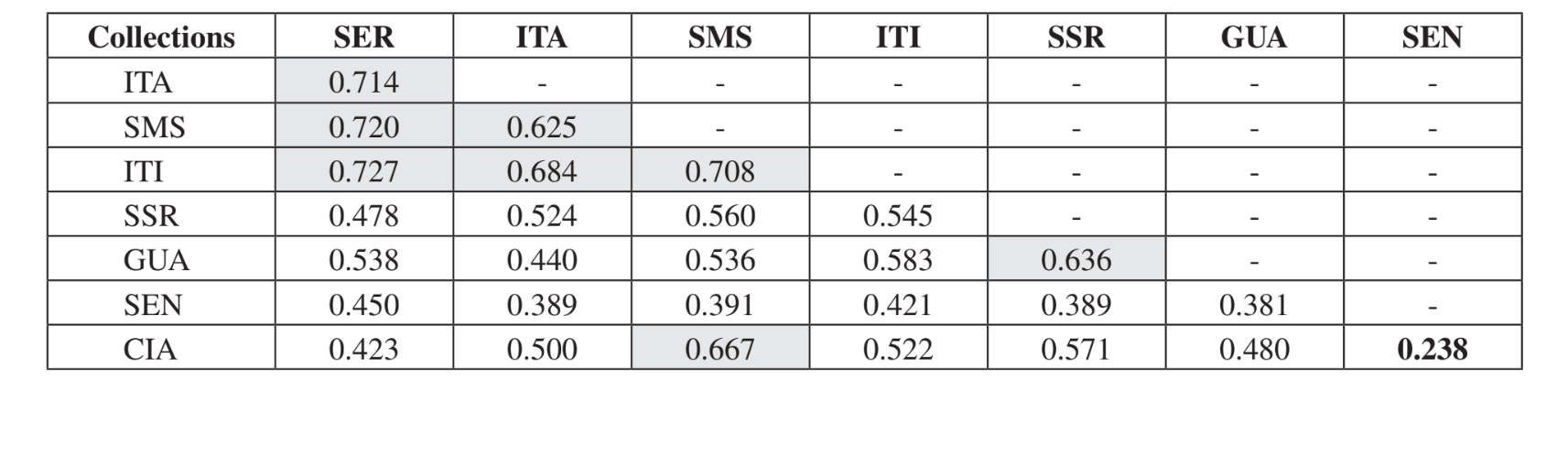
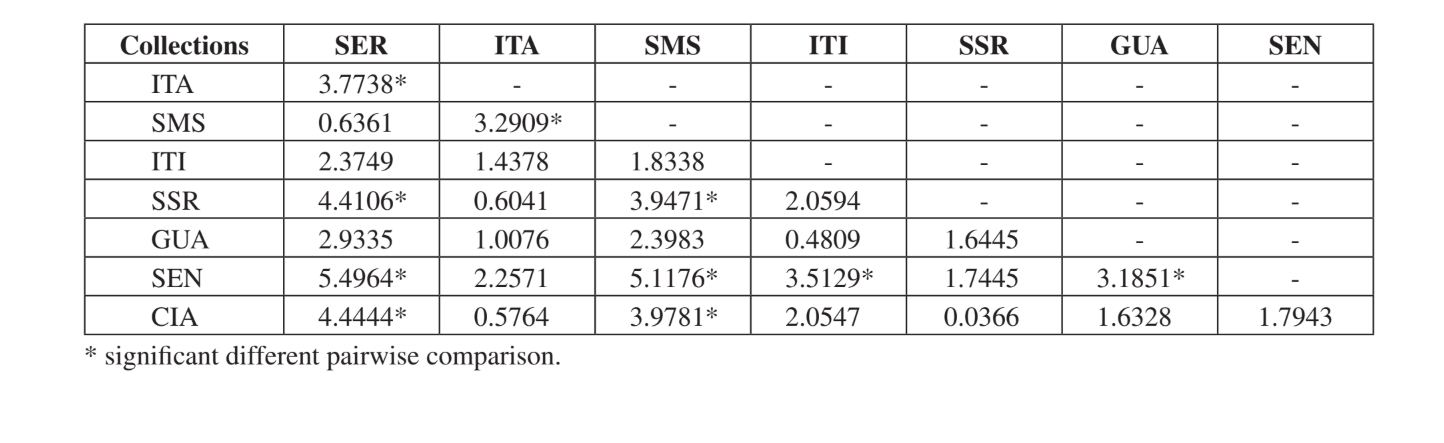
La prueba de Kruskal-Wallis, ajustada por empates, mostró que las comunidades son significativamente diferentes cuando se realizó una comparación general (H = 24.24; d.f. = 7; p = 0.001). Las comparaciones múltiples por pares de comunidades se presentan en la Tabla 5. Las comunidades de Serrana - SP, Santa Maria da Serra – SP y Sengés - PR fueron las más distintas cuando se compararon individualmente con las demás (4 comparaciones significativamente diferentes para cada una). Itirapina - SP y Canyon Guartelá - PR fueron las comunidades más similares cuando se compararon con cada comunidad muestreada (solo una diferencia significativa para cada una). Todas estas comparaciones (índice de Jaccard, análisis de clúster y comparaciones múltiples por pares) mostraron que ocurrió una mayor similitud entre las comunidades del Estado de São Paulo, aunque no se encontró una correlación significativa positiva en la prueba de Mantel que comparaba el índice de Jaccard y las distancias geográficas (r = – 0.3235; p = 0.0839).
La encuesta de composición de la comunidad es un paso importante para varios estudios en una amplia gama de campos, como la genética, la ecología y la evolución. Este trabajo añade nuevo conocimiento sobre la composición y diversidad de la comunidad de Drosophila en áreas xerofíticas que forman islas continentales en la región neotropical. Este enfoque es importante para futuros estudios en cualquier campo que considere a Drosophila como modelo.
Autores: Mateus, R. P., Buschini, M. L. T. y Sene, F. M
Referencias:
- Ab’saber, A. N., 1977, Espacios ocupados por la expansión de los climas secos de América del Sur, por ocasión de los períodos glaciares cuaternarios. Paleoclimas, 3: 1-19.
- Amorin, D. S. & Pires, M. R. S., 1996, Biogeografía neotropical y método para la estimación máxima de biodiversidad. En: C. E. M. Bicudo & N. A. Menezes (eds.), Biodiversidad en Brasil, un primer enfoque, CNPq, São Paulo.
- Bächli, G., 1998, Familia Drosophilidae. En: L. Papp & B. Darvas (eds.), Contribuciones a un Manual de Diptera Paleárticos. III. Brachycera Superior, Science Herald, Budapest.
- Belo, M. & Oliveira-Filho, J. J., 1978, Especies domésticas de Drosophila. V. Influencias de factores ambientales en el número de individuos capturados. Rev. Bras. Biol., 36: 903-909.
- Bigarella, J. J., Andrade-Lima, D. & Riehs, P. J., 1975, Consideraciones sobre los cambios paleoclimáticos en la distribución de algunas especies vegetales y animales en Brasil. An. Acad. Brasil. Ciênc., 41: 411-464.
- Bizzo, N. M. V. & Sene, F. M., 1982, Estudios sobre las poblaciones naturales de Drosophila de Peruíbe (SP), Brasil (Diptera, Drosophilidae). Rev. Bras. Biol., 42: 539-544.
- Burla, H., da Cunha, A. B., Cavalcanti, A. G. L., Pavan, C. & Dobzhansky, T., 1950, Densidad poblacional y tasas de dispersión en Drosophila willistoni brasileña. Ecology, 31: 393-404.
- Castle, W. E., 1906, Consanguinidad, cruce y esterilidad en Drosophila. Science, 23: 153.
- Dajoz, R., 1983, Ecología General. 471p. Editora Vozes, Petrópolis.
- Dobzhansky, T. & Pavan, C., 1943, Estudios sobre especies brasileñas de Drosophila. Bolm. Fac. Filos. Ciênc. S. Paulo, 36 (Biol. Geral, 4): 1-72.
- Dobzhansky, T. & Pavan, C., 1950, Variaciones locales y estacionales en las frecuencias relativas de especies de Drosophila en Brasil. J. Anim. Ecol., 19: 1-14.
- Duda, O., 1924a, Contribución a la sistemática de los Drosophilidae con especial consideración de las especies paleárticas y orientales (Diptera). Arch. Naturgesch., 90: 172-234.
- Duda, O., 1924b, Revisión de las especies europeas del género Drosophila Fallén (Diptera). Ent. Medd., 14: 246-313.
- Duda, O., 1925, Los Drosophilidae costarricenses del Museo Nacional Húngaro en Budapest. Annls hist.- nat. Mus. natn. hung., 22: 149-229.
- Duvernell, D. D. & Eanes, W. F., 2000, Genética poblacional molecular contrastante de cuatro hexoquinasas en Drosophila melanogaster, D. simulans y D. yakuba. Genetics, 156: 1191-1201.
- Grimaldi, D. A., 1990, Una clasificación filogenética revisada de los géneros en los Drosophilidae (Diptera). Bull. Am. Mus. nat. Hist., 197: 1-139.
- Klaczko, L. B., Powell, J. R. & Taylor, C. E., 1983, Drosophila cebos y levaduras: especies atraídas. Oecologia, 59: 411-413.
- Kratz, F. L., Pinto, L. G., Brandão, D. & Faria, L. G., 1982, Altura de vuelo y el patrón de distribución espacial en Drosophila. Ciênc. Cult., 34: 203-209.
- Medeiros, H. F., 2000, Asambleas de Especies de Drosophila (Diptera; Drosophilidae) y Efectos de Cursos de Agua sobre sus Distribuciones en dos Matas de São Paulo, 98p. Disertación (Maestría) – Instituto de Biología IB/UNICAMP, Campinas.
- Medeiros, H. F. & Klaczko, L. B., 2004, ¿Cuántas especies de Drosophila (Diptera, Drosophilidae) quedan por describir en los bosques de São Paulo, Brasil? Listas de especies de tres remanentes forestales. Biota Neotropica, 4: 1-12.
- Miller, M. P., 1997, Herramientas para análisis genéticos poblacionales - TFPGA - 1.3: Un programa de Windows para el análisis de datos genéticos alozímicos y moleculares. Software informático distribuido por el autor.
- Moraes, E. M., Mateus, R. P. & Sene, F. M., 2000, Ocurrencia y fluctuación en el tamaño poblacional de una especie de Drosophilidae recientemente introducida en Brasil. En: RESÚMENES DEL XXI CONGRESO INTERNACIONAL DE ENTOMOLOGÍA, LIBRO I. Actas..., 271p.
- Mourão, C. A., Gallo, A. J. & Bicudo, H. E. M. C., 1965, Sobre la sistemática de Drosophila en Brasil, con la descripción de D. mendeli sp.n. y “Relación de Especies Brasileñas del Género Drosophila”. Ciênc. Cult., 17: 577-585.
- Pavan, C., 1950, Especies Brasileñas de Drosophila. II. Bolm. Fac. Filos. Ciênc. S. Paulo, 111 (Biol. Geral, 8): 1-37.
- Pavan, C., 1959, Relaciones entre poblaciones naturales de Drosophila y el medio ambiente. Bolm. Fac. Filos. Ciênc. S. Paulo, 221 (Biol. Geral, 11): 1-81.
- Pavan, C. & da Cunha, A. B., 1947, Especies Brasileñas de Drosophila. Bolm. Fac. Filos. Ciênc. S. Paulo, 86 (Biol. Geral, 7): 3-47.
- Pavan, C., Dobzhansky, T. & Burla, H., 1950, Comportamiento diurno de algunas especies neotropicales de Drosophila. Ecology, 31: 36-43.
- Perondini, A. L. P., Sene, F. M. & Mori, L., 1979, El patrón y polimorfismo de algunas esterazas de Drosophila simulans en Brasil. Egypt. J. Genet. Cytol., 8: 263-268.
- Sene, F. M., Val, F. C., Vilela, C. R. & Pereira, M. A. Q. R., 1980, Datos preliminares sobre la distribución geográfica de especies de Drosophila dentro de los dominios morfoclimáticos de Brasil. Pap. Avul. Dep. Zool. Sec. Agric., 33: 315-326.
- Sene, F. M., Pereira, M. A. Q. R., Vilela, C. R. & Bizzo, N. M. V., 1981, Influencia de diferentes formas de colocar cebos para la recolección de moscas de Drosophila en tres ambientes naturales. D. I. S., 56: 118-121..
- Stalker, H. D., 1976, Estudios cromosómicos en poblaciones silvestres de D. melanogaster. Genetics, 82: 323-347.
- Sturtevant, A. H., 1921, Las especies norteamericanas de Drosophila. Carnegie Inst. Wash. Publ., 301: 1-150.
- Sturtevant, A. H., 1939, Sobre la subdivisión del género Drosophila. Proc. Natl. Acad. Sci. USA, 3: 137-141.
- Sturtevant, A. H., 1942, La clasificación del género Drosophila, con descripciones de nueve nuevas especies. Univ. Texas Publ., 4213: 6-51.
- Tidon-Sklorz, R. & Sene, F. M., 1992, Distribución vertical y temporal de especies de Drosophila (Diptera, Drosophilidae) en un área boscosa en el estado de São Paulo, Brasil. Rev. Bras. Biol., 52: 311-317.
- Tidon-Sklorz, R. & Sene, F. M., 1995, Fauna de Drosophila (Diptera, Drosophilidae) en el área norte de la “Cadeia do Espinhaço”, Estados de Minas Gerais y Bahía, Brasil: Aspectos biogeográficos y ecológicos. Iheringia, Sér. Zool., 78: 85-94.
- Tidon-Sklorz, R. & Sene, F.M., 1999, Drosophila. En: C. R. F. Brandão & E. M. Cancello (eds), Invertebrados Terrestres. Vol.V. Biodiversidad del Estado de São Paulo. Síntesis del Conocimiento al Final del Siglo XX, FAPESP, São Paulo.
- Tidon-Sklorz, R. & Sene, F. M., 2001, Dos nuevas especies del conjunto de hermanos de Drosophila serido (Diptera, Drosophilidae). Iheringia, Sér. Zool., 90: 141-146.
- Val, F. C. & Kaneshiro, K. Y., 1988, Drosophilidae (Diptera) de la Estação Biológica de Boracéia, en la cordillera costera del Estado de São Paulo, Brasil: Distribución geográfica. En: P. E. Vanzolini & W. R. Heyer (eds.), Actas de un Taller sobre Patrones de Distribución Neotropicales, Academia Brasileira de Ciências, Río de Janeiro.
- Val, F. C. & Sene, F. M., 1980, Una especie de Drosophila recién introducida en Brasil. Pap. Avul. Dep. Zool. Sec. Agric., 33: 293-298.
- Val, F. C., Vilela, C. R. & Marques, M. D., 1981, Drosophilidae de la Región Neotropical. En: M. Ashburner, H. L. Carson & J. N. Thompson (eds.), La genética y biología de Drosophila, Academic Press, Londres.
- Vanzolini, P. E., 1981, Un enfoque cuasi-histórico sobre la historia natural de la diferenciación de los reptiles en aislamientos geográficos tropicales. Pap. Avul. Dep. Zool. Sec. Agric., 34: 189-204
- Vermeij, G. J., 1996, Una agenda para la biología de invasiones. Biol. Conserv., 78: 3-9.
- Vilela, C. R., 1983, Una revisión del grupo de especies Drosophila repleta (Diptera, Drosophilidae). Rev. Bras. Entomol., 27: 114.
- Vilela, C. R., 1999, ¿Está Zaprionus indianus Gupta, 1970, (Diptera, Drosophilidae) colonizando actualmente la región neotropical? D. I. S., 82: 37-39.
- Vilela, C. R., Pereira, M. A. Q. R. & Sene, F. M., 1983, Datos preliminares sobre la distribución geográfica de especies de Drosophila dentro de los dominios morfoclimáticos de Brasil. II. El grupo repleta. Ciênc. Cult., 35: 66-70.
- Zar, J. H., 1999, Comparaciones Múltiples. En: Análisis Bioestadístico. 4ª ed., Prentice Hall Inc., Nueva Jersey.